
161 Years Ago
In the centuries old St. Thomas Abbey– in the garden and greenhouse– Gregor Mendel tended, crossed, and tracked some 10,000 pea plants (Pisum sativum) through several generations over 7 years. The greenhouse was warm compared to the monastery where his chambers were. The monastery was constantly damp and cool, originally built as a fortress in the 14th century, the Augustinian Monks were relatively new residents. Mendel was curious about how the world worked– he was into science and had broad interests– though he may not have used that term– the word science was coined and defined in England in 1834.
Mendel and the peas he planted, crossed, and counted started a revolution. It was quiet, at first, though Mendel knew he’d found something significant.
Peas were the second organism he studied in his experiments on trait inheritance. He started with mice. Those experiments were banned by Abbot Knapp in a compromise with the meddlesome local bishop, who felt the rats were filthy, that the work Mendel was doing was morally repugnant/against God’s will, and their litter smelled bad (The bishop was likely right on this last count).
Mendel and the peas he planted, crossed, and counted started a revolution. It was quiet, at first, though Mendel knew he’d found something significant. The degree of that significance was likely not clear to him at the time, however. 150 years ago, he presented and published it in a local journal and was then “lost” to the world. It’s more accurate to say it just didn’t spark anything in anyone until the right eyes and brain hit upon it. 34 years later, 4 brains rediscovered his work.
Activating Mendel’s Ideas
At the turn of the 20th century, having been cited by 3 other scientists also interested in inheritance. One of them, Hugo de Vries, was a student of Julius Von Sachs, who I wrote about a few months ago. William Bateson, a zoologist seeking a mechanistic explanation for Darwin’s natural selection, having seen Mendel’s paper cited several times by his continental colleagues, read Mendel’s paper on a train ride to giving a lecture. He tossed out what he was originally going to talk about and focused on Mendel’s ideas (Henig, 2001). With that, Mendel’s results were rediscovered, eventually dovetailing with natural selection, the structure of DNA, and the discovery of genes as physical objects made up of specific sequences of Adenine (A), Thiamine (T), Cytosine (C), and Guanine (G).
Mendel was intelligent. He was also part of an order of Augustinian Monks that encouraged the pursuit of knowledge. Mendel grew up relatively poor. His parents were farmers and Mendel did help tend fruit trees on the family farm. He was clearly a bright student, and qualified for school, university, and beyond (and thanks to family, he was able to finance his studies during a hard time when he may well not have been able to afford to continue onto university). Mendel also studied mathematics and physics. He pursued his curiosity in astronomy, meteorology, and what he is now best known for: genetics (not a term in use when Mendel did his work). Once done crossing peas, Mendel moved onto other pursuits as well as getting promoted to Abbot himself, leaving a lot less time for him to pursue research.

Despite being brief, Mendel’s foray into the life sciences was influential. And if Darwin had known about Mendel’s work (Mendel did send it to Darwin who never opened it), it may have answered some of his unknowns about his theory of natural selection that still lacked a clear method for “descent with modification” generation to generation. It would take until well into the 20th century for Mendel’s discrete factors of inheritance to be mapped onto DNA, to learn just what those heritable packets of information were, and the physical structure of genes– encoding the blueprint for an organism. Barbara McClintock showed in her observations of maize cells that chromosomes made up of DNA underwent physical changes during reproductive cell cycles.
Mendel worked with “true breeding” lines that had a consistent look- or phenotype as we’d say today– generation after generation (we’d also call them inbred lines). The first few years of his work were ensuring his starting material was standardized. Tall and dwarf, round and wrinkled, white and purple flowers– 7 traits in all that were known to vary- seemingly simply (Noell Ellis et al., 2011; Ross and Reid, 2011). Then he started crossing. Tall x dwarf. Round x wrinkled, etc. and he watched so called “recessive” traits disappear in the first generation after the cross only to re-appear . And then observed how two traits sorted independently of one another. Crossing round green pod plants with wrinkled yellow ones, for instance.
Using Mendelian Genetics

An extreme case of Mendelian-style crosses was carried out by George Beadle who analyzed 50,000 some progeny of a cross between maize and its wild ancestor, teosinte. Based on how the plants looked— maize-like, or teosinte-like and everything in between, he concluded that 4-5 genetic differences accounted for most of the differences between the two plants because around 1 in 500 2nd generation segregants after the initial cross looked like teosinte or maize alone (Doebley, 2001). Indeed, genes located at specific places in the maize genome, have been found that have “maize” versions and “teosinte” versions and depending on which versions a plant has, it will be more maize-like or teosinte-like (Doebley, 2004).
It’s easy to see Mendel’s work as underscoring how genes determine everything about how an organism turns out. Indeed, Mendel’s biggest legacy is probably that traits an organism has are underlain by discrete factors that we now understand as genes located at specific places along a chromosome– that get “expressed” (transcribed) into RNA and then many of those RNAs are further translated into proteins (some RNA molecules and proteins do the heavy lifting/work of carrying out functions of a cell/organism). However, Mendel studied what scientists now call “genes of major effect”. Most genomes of complex multicellular organisms (even some single cell organisms) have between 15-40,000 genes (bacteria/archaea generally have from 500-5,000). Most genes contribute in small ways to complex traits– like building a flagellum or eye. Often, in the plant world at least, genes come in families with overlapping/redundant functions– whole genome duplications are not uncommon in many plant’s evolutionary pasts. Often, the nature of a recessive mutation is the loss of a gene’s function. If one of a family is lost, the effect may not be apparent. The products of genes, RNA or proteins, also work as networks that are increasingly studied by life scientists.
One of Mendel’s pea traits, wrinkled peas, is thought to be due to the loss of function of a starch branching enzyme (Bhattacharyya et al., 1990). Starch is a branched polymer– like a tree– of sugar molecules, a way plants store sugars made via photosynthesis (the wrinkled peas are sweeter because the sugars aren’t branched). Genes come in versions, called alleles. In a simple case, a gene might have two versions in a population; one functional, one not. In reality, mutations, insertions, or differences in regulation can all exist in myriad forms, many alleles exist. Measuring the frequency and number of alleles of one gene in a population is one part of determining genetic diversity (and then multiply that by all the variation in all the genes of the genome). Of course, any individual only has a maximum of two alleles per gene.
Genes on the same chromosome often move through generations together; they are linked. The closer together two genes are, the more likely those versions of genes are to stay together when passed onto the next generation. During meiosis- the cell division process involved in reproduction- recombination can occur between similar chromosomes that have the most similar gene order on them. In eukaryotes, organisms with nuclei like humans and plants, two copies of every gene exist– one from “mom” and one from “dad”. If “mom” and “dad” have different alleles of a gene, those can swap places in a process called crossing over– thus shuffling combinations of genes on chromosomes (this is largely a random process, but the closer two genes are to one another, the less likely a recombination event in the space between them is).
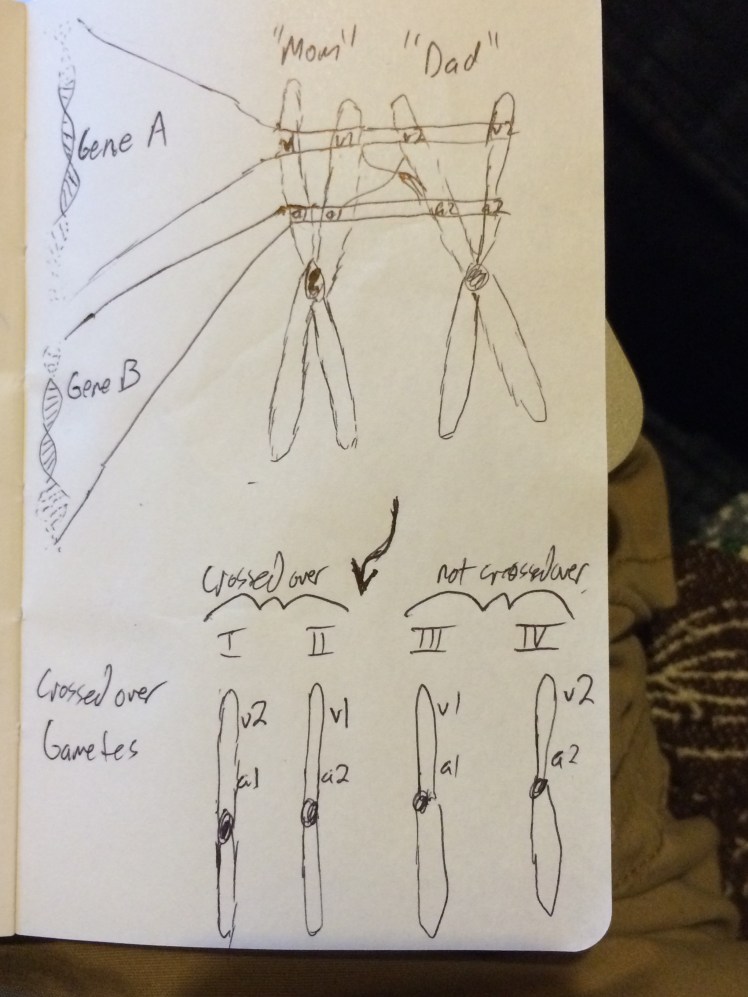
Mendel knew nothing of recombination, linkage, or crossing over. His “dominant”/”recessive” paradigm exists precisely because peas have two copies of their genome– one from each parent. If one copy is a functional gene and the other non-functional, the functional one is the “dominant” trait and seen in that first generation after the cross– round peas, for instance. The “recessive” trait can then segregate out again in the next generation because in ¼ of the cases, two “recessive”- nonfunctional copies of the gene can be inherited, revealing that phenotype, while ¾ will show the “dominant” phenotype. Mendel didn’t study any linked traits that we know of, and that is why his law of independent assortment when studying two traits at once– crossing wrinkled purple-flowered peas with round white ones, for instance– works (Ellis et al., 2011; Ross and Reid, 2011). That would have complicated his analysis, but it was precisely observations of non-Mendelian patterns of trait inheritance that led to the discovery of linked genes and the first genetic maps in the early 1900s out of Thomas Hunt Morgan’s lab. Part of Mendel’s insight was to bring probability to biology, something that continue on with Morgan’s work.
Mutating Mendel’s Ideas
In the 1930s and 40s, and 50s, inheritance and DNA became the explanation for how traits evolved and were selected for or against given the environment. In the 1960s and 70s and through today, the definition of “genes” has been honed and revised and is still complicated to define. It is also clear that the environment matters a lot in how genes express themselves. Organismal development and genetics also became integrated (Prunet and Meyerowitz, 2016). All of this is a growing inheritance of Mendel’s investigations into pea plants.

“Phenomics” is the study of all the possible phenotypes (physical traits) a plant of a given genotype (what alleles an organism has) might adopt (or the phenotypes of many different genotypes in a given environment). Genotypes are a lot easier to determine these days as gene sequencing technologies have advanced and gotten cheaper (the computing power/storage and learning analytic methods are the main barriers to large scale genotypic analyses now). Phenomics is a frontier in plant biology– a phenotype can be visible, like a leaf shape or down to the expression pattern of a gene in a cell– it has a lot more potential dimensions than a genotype. Plants are good for phenomic and gene by environment interaction studies (e.g. nature vs. nurture, though always it is some combination of both) because the environment can be kept more constant than for animals (that tend to move around). And the things we learn about how a plant’s genotype– its genes– interact with its environment can be applied to animals and other living things as well.

Mendel may have been overemphasized in genetics in the past 150 years. Biology and inheritance are more complex than just one gene for one trait most of the time. Even a simple “dominant” and “recessive” is not that common, but it was not a bad place to start. Mendel sketched out a rough outline of how inheritance worked through careful observation and mathematics. George Beadle is even more famous for his “one gene, one protein” hypothesis than his maize experiment outlined above (which is partly correct, but also more complex than that simple statement). Environmental factors also play a role in how genes wind up expressing themselves(get swtiched on or off), too and shape the organism as much as the specific combination of genes they have.
Genes aren’t the only way have to define our identity as science writer Diana Crow explored in a 3-part series on her blog based on a conference she attended (part 1, part 2, and part 3). It’s well worth the read. As our genomic databases have grown, it is clearly not sufficient to build identity upon– and the way genetics has been used (& still is in some cases), can be problematic– eugenics being a prominent example of how genes-as-destiny can go really wrong. Having the genome sequence of one organism is far from sufficient to fully understanding how it works since most living things have lots of other living things that are also a part of it in the form of microbes. A plant genome plus its microbiome from both leaf and soil environments are one example of a “holo-organism”– where the plant and microbes work together to generate the full plant behavior/growth characteristics. Pea plants, for example, are legumes, that have an important symbiosis with a bacteria in their roots enbling them to fix nitrogen. Genomes (expressing their contained genes) interact in complex ways and even transfer DNA between each other sometimes– it’s really complicated. This form of diversity is what we call “nature”. The living world that we all exist in and are connected to in more ways that we are consciously aware of. Though fictional, the TV show Orphan Black underscores the complexity of biology. The clones on the show are all slightly different (radically so personality-wise) despite having identical genomes.
Though it was slow, Mendel’s insights kicked off a revolution in genetics (even though many people had proposed ideas about how traits passed onto other generations before him). Along with other scientists, writers, philosophers, and teachers of his day, life science was able to be quantitatively studied, not just described (both things are valuable in science– but the former is far more emphasized in most fields of life science today). Mendel’s ideas still inform genetics today. And it all started with a bright Augustinian Monk growing peas in his greenhouse and garden.
References
Beans, C. Four Ways Inheritance is More Complex than Mendel Knew. 2016. NIGMS Biobeat. https://biobeat.nigms.nih.gov/2016/03/four-ways-inheritance-is-more-complex-than-mendel-knew/
Bhattacharyya MK, Smith, AM, Noel Ellis, TH, Hedley, C, Martin, C. The Wrinkled-Seed Character of Pea Described by Mendel is Caused by a Transposon-like Insertion in a Gene Encoding Starch-Branching Enzyme. 1990. Cell. 60: 115.
Diana Crow’s blog, 2016:
http://dianacrowscience.com/molecularization-indentity-3/
http://dianacrowscience.com/hybrid-problems/
http://dianacrowscience.com/perceive-identify-regulate-racist-science/
Doebley, J. George Beadle’s Other Hypothesis: One Gene, One Trait. 2001. Genetics 158: 487. http://www.genetics.org/content/158/2/487
Doebley, J. The Genetics of Maize Evolution. 2004. Ann. Rev. Genetics 38: 37-59http://teosinte.wisc.edu/pdfs/DoebleyAnnRev2004.pdf
Fleischman, John. The “Lost” Paper of Gregor Mendel, the Patron Saint of Low Impact Scientific Publishing. 2016, March 10. ASCB blog: http://www.ascb.org/lost-paper-gregor-mendel-patron-saint-low-impact-scientific-publishing/
Henig, Robin Marantz. The Monk in The Garden: The Lost and Found Genius of Gregor Mendel, The Father of Genetics. 2001. Mariner Books. ISBN-13: 978-0618127412
Noel Ellis TH, Hofer JMI, Timmerman-Vaughn GM, Coyne, CJ, Hellens, RP. Mendel, 150 Years On. 2011. Trends in Plant Science 16: 590. Doi: 10.1016/j.tplants.2011.06.006.
Nogoy, N. Complexities of Modern Phenomics. 2016 Biomed Central Giga Blog.
Prunet N, Meyerowitz EM. Genetics and Plant Development. 2016. C.R. Biologies In Press. Doi: http://dx.doi.org/10.1016/j.crvi.2016.05.003
Py-Lieberman, Evolution World Tour: Mendel’s Garden, Czech Republic. 2012 Smithsonian Magazine.
Radick, D. Teach Students The Biology of Their Time. 2016. Nature News & Comment.
http://www.nature.com/news/teach-students-the-biology-of-their-time-1.19936?WT.mc_id=FBK_NatureNews
Reid JB, Ross JJ. Mendel’s Genes: Toward a Full Molecular Characterization. 2011. Genetics 189: 3-10.
John Innes Centre Historical Collections: http://collections.jic.ac.uk/category/history-of-genetics/ :
Mendel and The Culture of Commemoration. 2015 John Innes Centre Historical Collections. http://collections.jic.ac.uk/mendel-and-the-culture-of-commemoration/
Celebrating the History of Peas and The International Year of The Pulse. 2016. John Innes Centre Historical Collections.
Gregor Mendel: Planting the Seeds of Genetics. 2006. The Field Museum Archived Exhibit: http://archive.fieldmuseum.org/mendel/about.asp

This is a very nice article. I am a member of the New York Linnaean Society (http://linnaeannewyork.org/index.html nature and bird watching). I think Linnaeus’s ideas must have helped Mendel https://en.wikipedia.org/wiki/Carl_Linnaeus .
LikeLike
Thanks! Almost certainly Linnaeus’ ideas did inform Mendel a bit. Though there’s also TA Knight who in 1799 did similar experiments as Mendel with peas even and Mendel may have known about Knight’s protocols that inspired his own more rigorous experiments. Just the idea of an ordered way of organizing species is useful.
LikeLiked by 1 person
Interesting. Thanks.
LikeLike